- The paper demonstrates that autoencoders can reliably differentiate between Euclidean and graph-based representations in simulated place cell data.
- It employs a dynamic 7x7 maze with octagonal tiles to simulate rat navigation, incorporating Gaussian noise and distance metrics under both encoding hypotheses.
- It finds that autoencoders outperform UMAP in noisy conditions, suggesting improved fidelity in representing spatial geometry from neural recordings.
Summary of Simulated Place Cell Recordings and Geometric Representation Hypotheses
Introduction to Place Cell Encoding
The study investigates hippocampal place cells, which traditionally are understood to encode animal spatial locations in physical environments. Two predominant hypotheses are explored: Euclidean, where place cells represent actual physical locations, and graph-based, where the encoding reflects topological aspects of the environment important for task-solving. Such differentiation in place cell encoding becomes critical when animals navigate through complex mazes, suggesting possible divergence from purely Euclidean interpretations towards task-relevant topological mappings.
Methodology
Behavioral Simulation and Neural Encoding
The experiment employs a dynamic maze setup consisting of a 7x7 grid of octagonal tiles to simulate rat navigation. Rats seek designated goals in the maze, fostering neural recordings that reflect the spatial trajectory towards these goals. The firing rate model for neurons takes into account maximal firing rate and Gaussian noise, bound by distance metrics under Euclidean or graph-based assumptions. Maze configurations are altered, maintaining consistent topology while varying Euclidean positions to test both hypotheses (Figure 1).
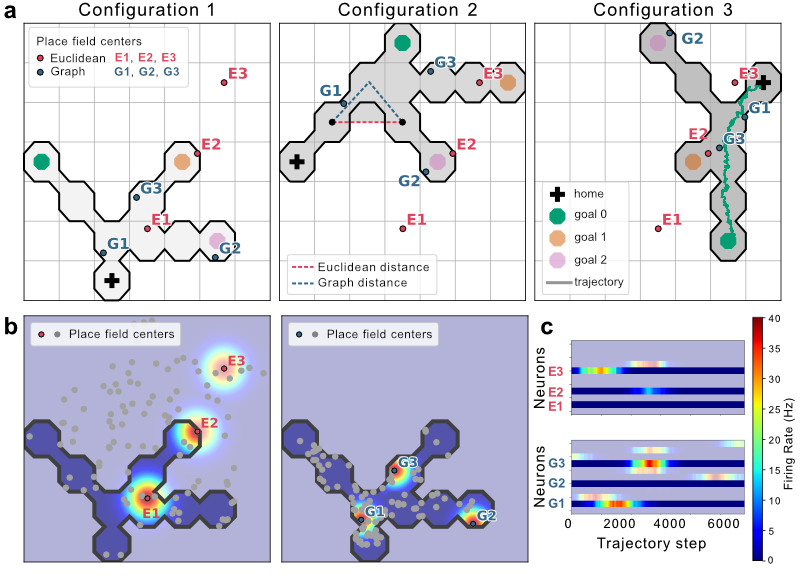
Figure 1: Maze configurations and simulated firing. (a) Three maze configurations showing place field centers of three example neurons for both hypotheses, Euclidean distance and dg(.)=2, and a simulated trajectory example.
Autoencoder Framework
Autoencoders (AEs) are utilized to analyze the simulated place cell data, learning embeddings within a 3D latent space that ideally reflects either hypothesis-driven spatial representation. The architecture is structured with equal depth encoder and decoder networks featuring leaky ReLU activations and linear or sigmoidal output layers, respectively. This approach emphasizes capturing complex latent relationships indicative of the encoded geometric structures.
Results
Latent Space Analysis through Autoencoders
Across multiple configurations, AEs demonstrate significant efficacy in capturing spatial geometry inherent to place cell data. Euclidean goal sharing across configurations manifests distinctly within latent space, suggesting fidelity to physical representation via AEs (Figure 2).
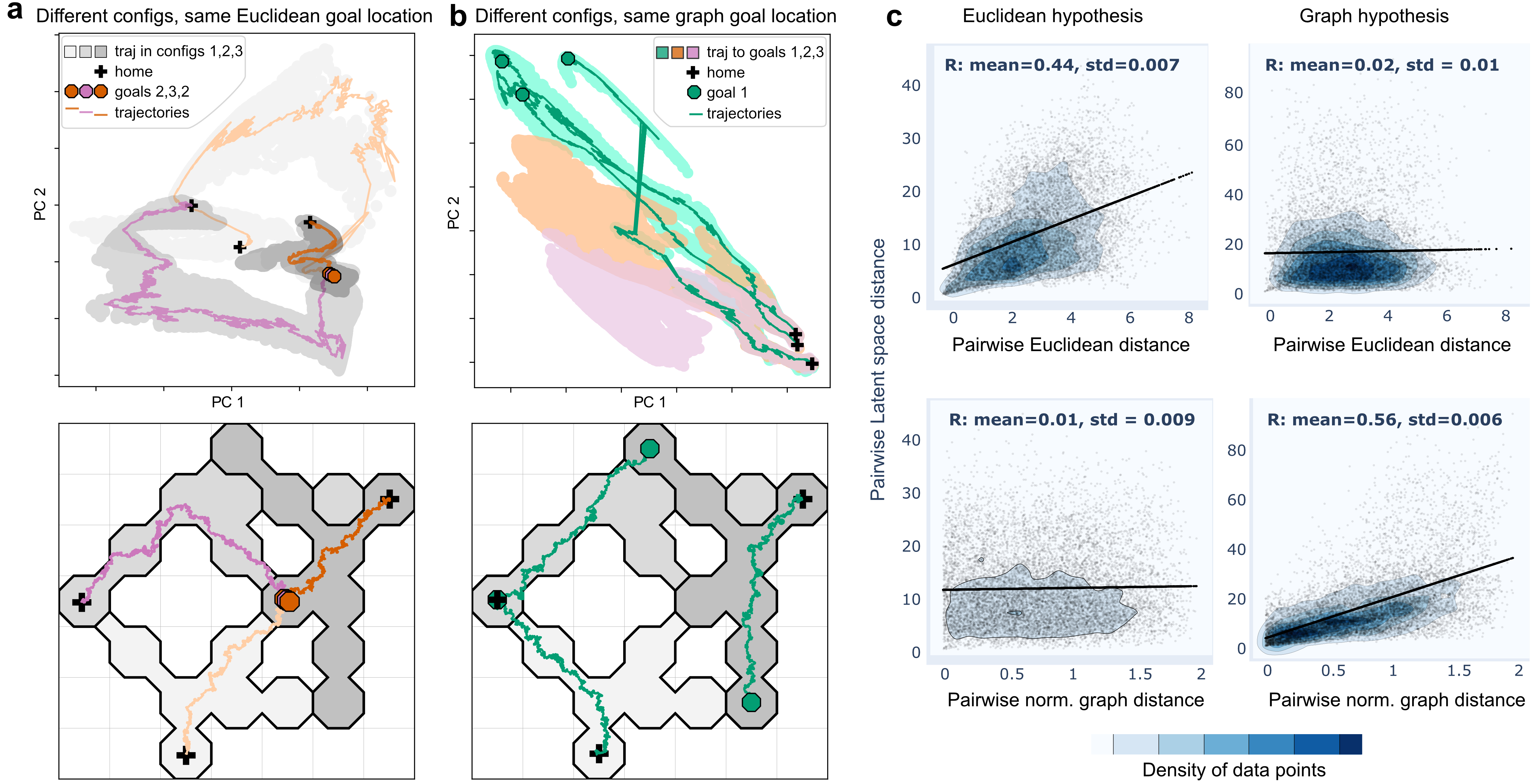
Figure 2: Latent space comparison (a) AE Latent states learned from the Euclidean dataset. (b) Same for the Graph dataset. (c) Comparison of latent space distances with physical and topological metrics.
Evaluation with UMAP and Comparative Approaches
While UMAP also embeds trajectories into latent dimensions, its robustness fluctuates under noisy conditions, unlike AEs which show sustained correlation integrity across metrics. This highlights AEs as particularly viable for latent space representation of neural data, especially under sparse encoding scenarios. (Figures 3 and 4)
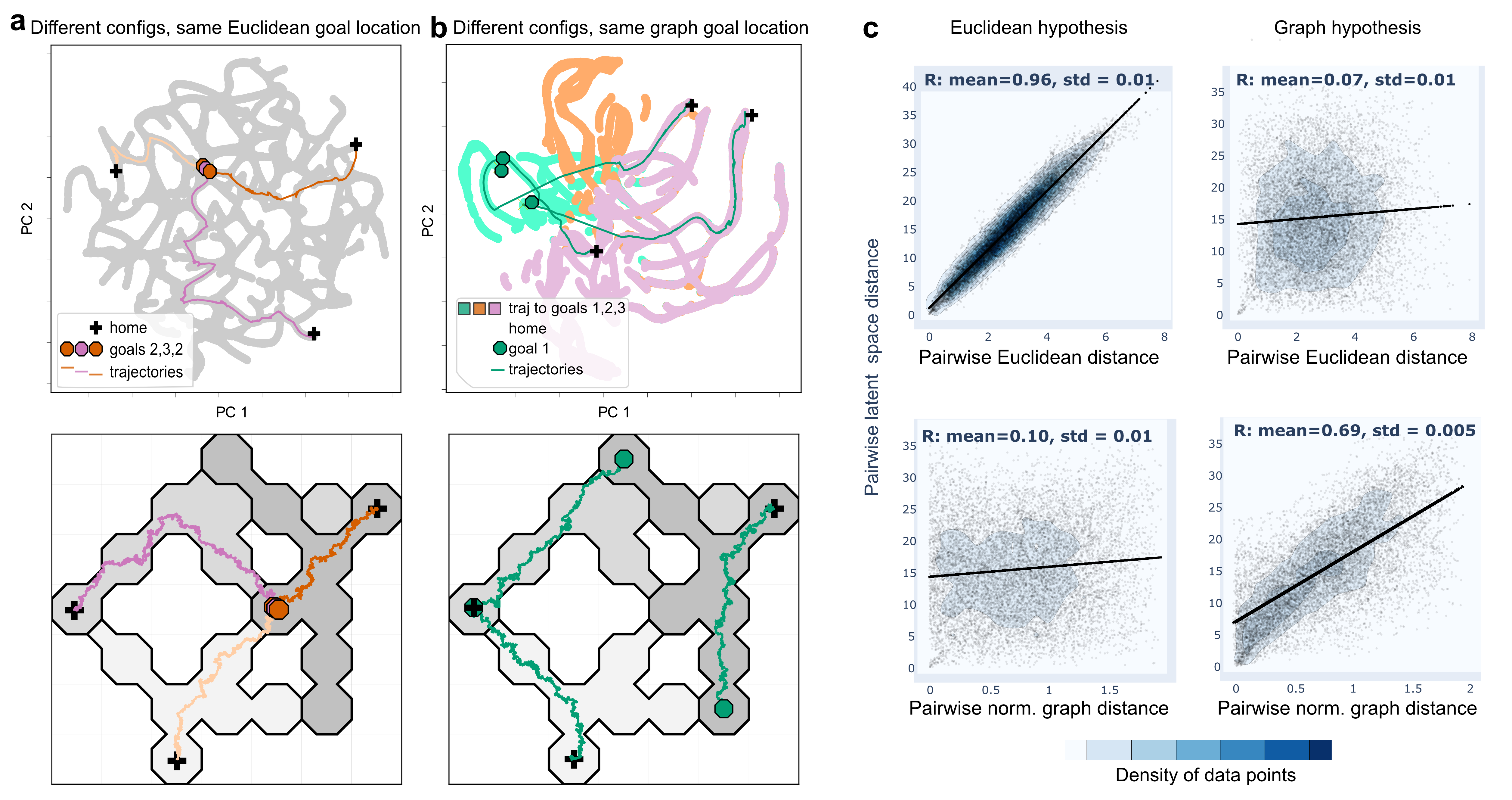
Figure 3: UMAP Latent space comparison (a) Latent states from the Euclidean dataset. (b) Same for the graph dataset.
Conclusion
The study delineates a framework where AEs and UMAP can discern between Euclidean and graph-based neural encoding in spatial tasks. The AE architecture outperforms in terms of noise resilience and representation clarity. Future research directions might involve comparative analysis across varied neural network configurations, enhancing task complexity, and incorporating dynamic neural encoding models to simulate real-world navigation.
Research advancements in this space have strong implications for understanding spatial cognition, especially how environments shape neural encoding within the hippocampus, which may inspire new methodologies in artificial intelligence navigation systems on mazes and graph-based paths.

